
Os fungos são classificados num reino separado das plantas, animais e bactérias. Uma grande diferença é o facto de as células dos fungos terem paredes celulares que contêm quitina, ao contrário das células vegetais, que contêm celulose. Estas e outras diferenças mostram que os fungos formam um só grupo de organismos relacionados entre si, denominado Eumycota (fungos verdadeiros ou Eumycetes), e que partilham um ancestral comum (um grupo monofilético). Este grupo de fungos é distinto dos estruturalmente similares Myxomycetes (agora classificados em Myxogastria) e Oomycetes. A disciplina da biologia dedicada ao estudo dos fungos é a micologia, muitas vezes vista como um ramo da botânica, mesmo apesar de os estudos genéticos terem mostrado que os fungos estão mais próximos dos animais do que das plantas.
Abundantes em todo mundo, a maioria dos fungos é inconspícua devido ao pequeno tamanho das sua estruturas, e pelos seus modos de vida crípticos no solo, na matéria morta, e como simbiontes ou parasitas de plantas, animais, e outros fungos. Podem tornar-se notados quando frutificam, seja como cogumelos ou como bolores. Os fungos desempenham um papel essencial na decomposição da matéria orgânica e têm papéis fundamentais nas trocas e ciclos de nutrientes. São desde há muito tempo utilizados como uma fonte direta de alimentação, como no caso dos cogumelos e trufas, como agentes levedantes no pão, e na fermentação de vários produtos alimentares, como o vinho, a cerveja, e o molho de soja. Desde a década de 1940, os fungos são usados na produção de antibióticos, e, mais recentemente, várias enzimas produzidas por fungos são usadas industrialmente e em detergentes. São também usados como agentes biológicos no controlo de ervas daninhas e pragas agrícolas. Muitas espécies produzem compostos bioativos chamados micotoxinas, como alcaloides e policetídeos, que são tóxicos para animais e humanos. As estruturas frutíferas de algumas espécies contêm compostos psicotrópicos, que são consumidos recreativamente ou em cerimónias espirituais tradicionais. Os fungos podem decompor materiais artificiais e construções, e tornar-se patogénicos para animais e humanos. As perdas nas colheitas devidas a doenças causadas por fungos ou à deterioração de alimentos pode ter um impacto significativo no fornecimento de alimentos e nas economias locais.
O reino dos fungos abrange uma enorme diversidade e táxons, com ecologias, estratégias de ciclos de vida e morfologias variadas, que vão desde os quitrídios aquáticos unicelulares aos grandes cogumelos. Contudo, pouco se sabe da verdadeira biodiversidade do reino Fungi, que se estima incluir 1,5 milhões de espécies, com apenas cerca de 5% destas formalmente classificadas. Desde os trabalhos taxonómicos pioneiros dos séculos XVII e XVIII efetuados por Lineu, Christiaan Hendrik Persoon, e Elias Magnus Fries, os fungos são classificados segundo a sua morfologia (i.e. caraterísticas como a cor do esporo ou caraterísticas microscópicas) ou segundo a sua fisiologia. Os avanços na genética molecular abriram o caminho à inclusão da análise de ADN na taxonomia, o que desafiou por vezes os antigos agrupamentos baseados na morfologia e outros traços. Estudos filogenéticos publicados no último decénio têm ajudado a modificar a classificação do reino Fungi, o qual está dividido em um sub-reino, sete filos e dez subfilos.
Etimologia
Características
A palavra portuguesa fungo deriva do termo latino fungus (cogumelo), usado nos escritos de Horácio e Plínio, o Velho.3 Por seu lado, fungus é derivado do grego sphongos/σφογγος ("esponja"), que se refere às estruturas e morfologia macroscópicas dos cogumelos e bolores. O termomicologia, derivado do grego mykes/μύκης (cogumelo) e logos/λόγος (discurso),4 para denotar o estudo científico dos fungos, terá sido usado pela primeira vez em 1836, pelo naturalista inglês Miles Joseph Berkeley na obra The English Flora of Sir James Edward Smith, Vol. 5.5
Antes da introdução dos métodos moleculares de análise filogenética, os taxonomistas consideravam que os fungos eram membros do reino Plantae devido a semelhanças nos seus modos de vida: tanto os fungos como as plantas são na sua maioria imóveis, e apresentam semelhanças na morfologia geral e no habitat em que se desenvolvem. Tal como as plantas, muitas vezes os fungos crescem no solo, e no caso dos cogumelos formam corpos frutíferos conspícuos, que por vezes se assemelham a plantas como os musgos. Os fungos são agora considerados um reino separado, distintos das plantas e animais, dos quais parecem ter divergido há cerca de mil milhões de anos.6 7 Algumas caraterísticas morfológicas, bioquímicas, e genéticas são partilhadas com outros organismos, enquanto outras são exclusivas dos fungos, separando-os claramente dos outros reinos:
Caraterísticas partilhadas:
- Com os demais eucariotas: como nos restantes eucariotas, os núcleos das células dos fungos estão limitados por uma membrana e contêm cromossomas que contêm ADN com regiões não-codificantes chamadas intrões e regiões codificantes chamadas exões. Além disso, os fungos possuem organelos citoplasmáticos delimitados por membrana tais como mitocôndrias, membranas que contêm esterois, e ribossomas do tipo 80S.8 Têm um conjunto caraterístico de carboidratos e compostos armazenados solúveis, incluindo polióis (como manitol),dissacarídeos (como a trealose) e polissacarídeos (como o glicogénio, que também é encontrado em animais9 ).
- Com os animais: os fungos carecem de cloroplastos e são organismos heterotróficos, requerendo compostos orgânicos preformados como fontes de energia.10
- Com as plantas: os fungos possuem uma parede celular11 e vacúolos.12 Reproduzem-se por meios sexuados e assexuados, e tal como os grupos basais de plantas (como os fetos emusgos) produzem esporos. Tal como os musgos e algas, os fungos têm núcleos tipicamente haploides.13
- Com os euglenoides e bactérias: os fungos mais desenvolvidos, os euglenoides e algumas bactérias, produzem o aminoácido L-lisina em passos específicos de biossíntese, a via do alfa-aminoadipato.14 15
- As células da maioria dos fungos crescem como estruturas tubulares, alongadas e filamentosas designadas hifas. Estas podem conter múltiplos núcleos e crescer a partir das suas extremidades. Cada extremidade contém um conjunto de vesículas - estruturas celulares compostas por proteínas, lípidos e outras moléculas orgânicas - chamado Spitzenkörper.16 Tanto fungos como Oomycetes crescem como células hifais filamentosas.17 Em contraste, organismos de aspecto semelhante, como as algas verdes filamentosas, crescem por divisão celular repetida ao longo de uma cadeia de células.9
- Em comum com algumas espécies de plantas e animais, mais de 60 espécies de fungos apresentam bioluminescência.18
Caraterísticas únicas:
- Algumas espécies crescem como leveduras unicelulares que se reproduzem por gemulação ou por fissão binária. Os fungos dimórficos podem alternar entre uma fase de levedura e uma fase com hifas, em função das condições ambientais.19
- A parede celular dos fungos é composta por glicanos e quitina; enquanto os primeiros são também encontrados em plantas e a última no exosqueleto dos artrópodes,20 21 os fungos são os únicos organismos que combinam estas duas moléculas estruturais na sua parede celular. Ao contrário das plantas e dos Oomycetes, as paredes celulares dos fungos não contêm celulose.22

A maioria dos fungos carece de um sistema eficiente para o transporte de água e nutrientes a longa distância, como o xilema e o floema de muitas plantas. Para ultrapassar estas limitações, alguns fungos, como os do género Armillaria, formam rizomorfos23 que são morfológica e funcionalmente semelhantes às raízes das plantas. Outra característica partilhada com as plantas consiste numa via bioquímica para a produção de terpenos que usaácido mevalónico e pirofosfato como precursores.24 Porém, as plantas têm uma via bioquímica para a produção de terpenos nos seus cloroplastos, uma estrutura que os fungos não possuem.25 Os fungos produzem vários metabolitos secundários que são estruturalmente semelhantes ou idênticos aos produzidos pelas plantas.24 Muitas das enzimas de plantas e fungos que produzem estes compostos diferem entre si na sequência de aminoácidos e outras características, o que indica origens e evolução separadas destas enzimas nos fungos e plantas.24 26
Diversidade

Os fungos têm uma distribuição mundial, e desenvolvem-se numa grande variedade de habitats, incluindo ambientes extremos como desertos ou áreas com elevadas concentrações de sais27 ou radiações ionizantes,28 bem como em sedimentos de mar profundo.29 Alguns podem sobreviver às intensas radiações ultravioleta e cósmica encontradas durante as viagens espaciais.30
A maioria desenvolve-se em ambientes terrestres, embora várias espécies vivam parcial ou totalmente em ambientes aquáticos, como o fungo quitrídioBatrachochytrium dendrobatidis, um parasita responsável pelo declínio global das populações de anfíbios. Este organismo passa parte do seu ciclo de vida na forma de um zoósporo móvel, o que lhe permite propulsar-se através da água e entrar no seu hóspede anfíbio.31 Outros exemplos de fungos aquáticos incluem aqueles que vivem em zonas hidrotermais dos oceanos.32
Estão descritas formalmente pelos taxonomistas cerca de 100 000 espécies de fungos,33 mas a biodiversidade global do reino dos fungos não é totalmente compreendida.34 Com base em observações do quociente entre o número de espécies de fungos e o número de espécies de plantas em ambientes selecionados, estima-se que o reino dos fungos contenha cerca de 1,5 milhões de espécies.35 Em termos históricos, em micologia, as espécies têm sido distinguidas por vários métodos e conceitos. A classificação baseada nas caraterísticas morfológicas, como o tamanho e forma dos esporos ou das estruturas frutíferas, tem dominado tradicionalmente a taxonomia dos fungos.36 As espécies podem também ser distinguidas pelas suas caraterísticas bioquímicas e fisiológicas, tais como a sua capacidade para metabolizar certos compostos bioquímicos, ou a sua reação a testes químicos. O conceito biológico de espécie discrimina as espécies com base na sua capacidade de acasalamento. A aplicação de ferramentas moleculares, como a sequenciação de ADN e a análise filogenética, no estudo da diversidade melhorou significativamente a resolução e aumentou a robustez das estimativas da diversidade genética nos vários grupos taxonómicos.37
Morfologia
Estruturas microscópicas

A maioria dos fungos desenvolve-se como hifas, que são estruturas filamentosas, cilíndricas, com dois a 10 µm de diâmetro e até vários centímetros de comprimento.
As hifas crescem nas suas extremidades (ápices); novas hifas formam-se tipicamente por meio da emergência de novas extremidades ao longo da hifa existente num processo designado “ramificação”, ou ocasionalmente por bifurcação de extremidades de uma hifa em crescimento, dando origem a duas hifas com crescimento paralelo.38 A combinação do crescimento apical com a ramificação/bifurcação conduz ao desenvolvimento de um micélio, uma rede interconectada de hifas.19 As hifas podem ser septadas ou cenocíticas: as hifas septadas são divididas em compartimentos separados por paredes transversais (paredes celulares internas, chamadas septos, que se formam perpendicularmente à parede celular, dando à hifa a sua forma), cada compartimento contendo um ou mais núcleos; as hifas cenocíticas não são compartimentadas.39 Os septos têm poros que permitem a passagem de citoplasma, organelos, e por vezes núcleos; um exemplo é o septo doliporo dos fungos do filo Basidiomycota.40 As hifas cenocíticas são essencialmente supercélulas multinucleadas.41
Muitas espécies desenvolveram estruturas hifais especializadas na absorção de nutrientes dos hospedeiros vivos; dois exemplos são os haustóriosnas espécies parasitas de plantas da maioria dos filos de fungos, e os arbúsculos de vários fungos micorrízicos, que penetram nas células do hospedeiro para consumir nutrientes.42
Embora os fungos sejam opistocontes – um agrupamento de organismos evolutivamente aparentados, caraterizados em termos gerais por possuírem um único flagelo posterior – todos os filos, exceto o dos quitrídios, perderam os seus flagelos posteriores.43 Os fungos são incomuns entre os eucariotas por terem uma parede celular que, além dos glicanos (p.e. β-1,3-glicano) e outros componentes típicos, contém também o biopolímero quitina.44
Estruturas macroscópicas

Os micélios dos fungos podem tornar-se visíveis a olho nu, por exemplo, em várias superfícies e substratos, tais como paredes húmidas e comida deteriorada, sendo vulgarmente chamados bolores. Os micélios desenvolvidos em meio de ágar sólido em placas de Petri de laboratório são usualmente designados colónias. Estas colónias podem apresentar formas e cores de crescimento (devido aos esporos ou a pigmentação) que podem ser usadas como caraterísticas de diagnóstico na identificação de espécies ou grupos.45 Algumas colónias individuais de fungos podem atingir dimensões e idades extraordinárias, como é o caso de uma colónia clonal de Armillaria ostoyae, que se estende por mais de 900 ha, com uma idade estimada em cerca de 9 000 anos.46
O apotécio – uma estrutura especializada importante na reprodução sexuada de Ascomycetes – é um corpo frutífero em forma de taça que contém ohiménio, uma camada de tecido contendo as células portadoras de esporos.47 Os corpos frutíferos dos basidiomicetes e de alguns ascomicetes podem, por vezes, atingir grandes dimensões, e muitos são bem conhecidos como cogumelos.
Crescimento e fisiologia
O crescimento dos fungos como hifas em substratos sólidos ou como células singulares em ambientes aquáticos, está adaptado para a extração eficiente de nutrientes, pois estas formas de crescimento têm uma razão entre a área superficial e o volume bastante alta.48 As hifas estão especificamente adaptadas ao crescimento sobre superfícies sólidas e à invasão de substratos e tecidos.49 Podem exercer grandes forças mecânicas penetrativas; por exemplo, o patógeno vegetal Magnaporthe grisea forma uma estrutura chamada apressório que evoluiu de forma a perfurar tecidos vegetais.50 A pressão gerada pelo apressório, dirigida contra a epiderme da planta, pode exceder os 8 MPa (80 bar).50 O fungo filamentoso Paecilomyces lilacinus, usa uma estrutura semelhante para penetrar os ovos de nemátodos.51

A pressão mecânica exercida pelo apressório é gerada a partir de processos fisiológicos que aumentam o turgor intracelular ao produzirem osmólitoscomo o glicerol.52 Adaptações morfológicas como estas são complementadas por enzimas hidrolíticas segregadas para o ambiente para a digestão de grandes moléculas orgânicas – como polissacarídeos, proteínas, lípidos, e outros substratos orgânicos – em moléculas menores que podem então ser absorvidas como nutrientes.53 54 55 A vasta maioria dos fungos filamentosos cresce de um modo polar – i.e., por extensão numa direção – por alongamento no ápice da hifa.56 Formas alternativas de crescimento dos fungos incluem a extensão intercalar (i.e. por expansão longitudinal de compartimentos hifais que estão abaixo do ápice), como é o caso em alguns fungos endófitos,57 ou o crescimento por expansão do volume durante o desenvolvimento dos estipes dos cogumelos e doutros grandes órgãos.58 O crescimento dos fungos como estruturas multicelulares consistindo de células somáticas e reprodutoras – uma característica que evoluiu de modo independente nos animais e plantas59 - tem várias funções, incluindo o desenvolvimento de corpos frutíferos para a disseminação dos esporos sexuais (ver acima) e de biofilmes para a colonização de substratos e comunicação intercelular.60
Tradicionalmente, os fungos são considerados heterotróficos, organismos que dependem exclusivamente do carbono fixado por outros organismos para o seu metabolismo. Os fungos desenvolveram um grau elevado de versatilidade metabólica, o que lhes permite utilizar uma variedade de substratos orgânicos para o seu crescimento, incluindo compostos simples como nitrato, amónia, acetato, ou etanol.61 62 Demonstrou-se para algumas espécies que o pigmento melanina pode ter um papel na extração de energia da radiação ionizante, como a radiação gama; porém, esta forma de crescimentoradiotrófico foi descrita apenas em algumas poucas espécies, os efeitos nas velocidades de crescimento são pequenos, e os processos biofísicos e bioquímicos subjacentes são desconhecidos.28 Os autores especulam que este processo pode ter semelhança com a fixação do dióxido de carbono vialuz visível, mas utilizando radiação ionizante como a fonte de energia.63
Reprodução

A reprodução dos fungos é complexa, refletindo as diferenças de modos de vida e da constituição genética existentes neste reino.64 Estima-se que um terço de todos os fungos pode reproduzir-se usando mais do que um modo de propagação; por exemplo, a reprodução pode ocorrer em dois estágios bem diferenciados no ciclo de vida de uma espécie, o teleomorfo e o anamorfo.65 As condições ambientais desencadeiam estados de desenvolvimento geneticamente determinados que conduzem à criação de estruturas especializadas para a reprodução sexuada ou assexuada. Estas estruturas auxiliam a reprodução ao dispersarem eficientemente esporos ou propágulos contendo esporos.
Reprodução assexuada
A reprodução assexuada por meio de esporos vegetativos (conídios) ou através da fragmentação do micélio é comum; ela mantém populações clonaisadaptadas a um nicho ecológico específico e permite uma dispersão mais rápida do que a reprodução sexuada.66 Os fungi imperfecti ouDeuteromycota (fungos que não apresentam estágio sexuado) incluem todas as espécies que não possuem um ciclo sexual observável.67
Reprodução sexuada
A reprodução sexuada com meiose existe em todos os filos de fungos, exceto Glomeromycota.68 Difere da reprodução sexuada de animais e plantas. Existem também diferenças entre grupos de fungos, as quais podem ser usadas para discriminar espécies em função de diferenças morfológicas nas estruturas sexuais e das estratégias de reprodução.69 70 Experiências de acasalamento entre isolados de fungos podem identificar espécies com base no conceito biológico de espécie.70 Os principais agrupamentos de fungos foram inicialmente delineados com base na morfologia das suas estruturas sexuais e esporos; por exemplo, as estruturas portadoras de esporos, ascos e basídios, podem ser usadas na identificação de ascomicetes e basidiomicetes, respetivamente. Algumas espécies permitem o acasalamento apenas entre indivíduos de tipo reprodutor oposto, enquanto noutras podem acasalar e reproduzir-se sexuadamente com qualquer outro indivíduo ou com eles mesmos. As primeiras dizem-se heterotálicas e as segundas homotálicas.71
A maioria dos fungos tem um estágio haploide e um estágio diploide nos seus ciclos de vida. Nos fungos de reprodução sexuada, os indivíduos compatíveis podem combinar-se fundindo as suas hifas numa rede interconetada; este processo, anastomose, é requerido para o início do ciclo sexual. Os ascomicetes e os basidiomicetes passam por um estágio dicariótico, no qual os núcleos herdados dos dois pais não se combinam imediatamente após a fusão celular, antes permanecendo separados nas células hifais (ver heterocariose).72
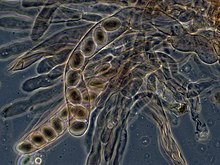
Nos ascomicetes, as hifas dicarióticas do himénio (a camada de tecido portador de esporos) formam um gancho caraterístico no septo hifal. Durante adivisão celular, a formação do gancho assegura a correta distribuição dos núcleos recém-divididos nos compartimentos hifais apical e basal. Forma-se então um asco, no qual ocorre cariogamia (fusão dos núcleos). Os ascos estão contidos num ascocarpo, ou corpo frutífero. A cariogamia nos ascos é imediatamente seguida de meiose e pela produção de ascósporos. Após a dispersão, os ascósporos podem germinar e formar um novo micélio haploide.73
A reprodução sexuada dos basidiomicetes é semelhante à dos ascomicetes. Hifas haploides compatíveis fundem-se para dar origem a um micélio dicariótico. Porém, a fase dicariótica é mais extensa nos basidiomicetes, estando muitas vezes presente também no micélio em crescimento vegetativo. Uma estrutura anatómica especializada, chamada fíbula, forma-se em cada septo hifal. Tal como com o gancho estruturalmente similar dos ascomicetes, a fíbula dos basidiomicetes é requerida para a transferência controlada de núcleos durante a divisão celular, para manter um estágio dicariótico com dois núcleos geneticamente diferentes em cada compartimento hifal.74 Forma-se um basidiocarpo, no qual estruturas em forma de bastão chamadas basídios geram basidiósporos haploides após cariogamia e meiose.75 Os basidiocarpos mais vulgarmente conhecidos são oscogumelos, mas também podem assumir outras formas (ver secção Morfologia).
Nos Glomeromycetes (antes Zygomycetes), as hifas haploides de dois indivíduos fundem-se, formando um gametângio, uma estrutura celular especializada que se torna uma célula produtora de gâmetas férteis. O gametângio evolui para um zigósporo, um esporo com parede espessa formado pela união de gâmetas. Quando o zigósporo germina, sofre meiose, gerando novas hifas haploides, as quais podem então formar esporangiósporos assexuados. Estes esporangiósporos permitem ao fungo dispersar-se rapidamente e germinar como micélios haploides geneticamente idênticos.76
Dispersão de esporos

Tanto os esporos assexuados como os sexuados (esporangiósporos) são frequentemente dispersos por ejeção forçada desde as suas estruturas reprodutoras. Esta ejeção garante a saída dos esporos das estruturas reprodutoras bem como a deslocação através do ar por grandes distâncias.
Mecanismos fisiológicos e mecânicos especializados, bem como as estruturas superficiais dos esporos (como as hidrofobinas), permitem a ejeção eficiente do esporo.77 Por exemplo, a estrutura das células portadoras de esporos de algumas espécies de ascomicetes é tal, que a acumulação desubstâncias que afetam o volume celular e o equilíbrio de fluidos, permite a descarga explosiva dos esporos no ar.78 A descarga forçada de esporos individuais, designados «balistósporos», envolve a formação de uma pequena gota de água (gota de Buller), que por contacto com o esporo leva à sua libertação com uma aceleração inicial superior a 10 000g;79 o resultado é o esporo ser ejetado a 0,01 – 0,02 cm, distância suficiente para que caia através das lamelas, ou poros, para o ar abaixo.80 Outros fungos, como os do género Lycoperdon, dependem de mecanismos alternativos para a libertação dos esporos, como forças mecânicas exteriores. O fungo ninho-de-pássaro usa a força das gotas de água em queda para libertar os esporos dos corpos frutíferos em forma de taça.81 Outra estratégia é observada em Phallaceae, um grupo de fungos com cores vivas e odor pútrido, que atraem insetos para dispersarem os seus esporos.82
Outros processos sexuados
Além da reprodução sexuada normal com meiose, certos fungos, como os dos géneros Penicillium e Aspergillus, podem trocar material genético por processos parassexuais, iniciados pela anastomose entre as hifas e plasmogamia das células dos fungos.83 A frequência e importância relativa dos eventos parassexuais não são claras, e podem ser menores que as dos outros processos sexuados. Sabe-se que tem um papel na hibridização intra-específica.84 e é provavelmente requerida para a hibridização entre espécies, a qual foi associada com os principais eventos na evolução dos fungos.85
0 comentários:
Postar um comentário